Кто станет магистром?..
Бакалавр – получивший бакалаврство, 1-ю степень в иноземных университетах.
Магистр – глава и старейшина некоторых военно-духовных орденов или братств;
получивший степень магистра, среднюю между докторской и кандидатской.Из Толкового словаря живого великорусского языка В.Даля
Подготовительный этап
Игра проводится по сценарию, сходному
со сценариями телеигр «Кто хочет стать
миллионером?» и «О, счастливчик!».
Учащимся заранее (за 2–3 урока) сообщается тема
игры, указывается материал, который надо
повторить, предоставляется необходимая
литература. Каждый участник игры должен
подготовить 4 карточки с буквами А, Б, В, Г
(рекомендуемый размер карточек: 15 х 15 см). Из
состава учащихся назначаются 2–3 помощника
ведущего (члены счетной комиссии), в роли
которого может выступить как сам учитель, так и
старшеклассник.
Класс заранее делится на 4–5 групп для проведения
отборочного тура (учитель доводит до их сведения,
что в спорных ситуациях ведущий будет
руководствоваться учебным рейтингом учащихся).
Правила игры
Игра состоит из двух туров:
отборочного и основного. Начинает игру ведущий.
В отборочном туре работа проводится в группах.
Помощники раздают в группы заранее
подготовленные листки, в которых надо расставить
буквы в правильной очередности. Та команда,
которая раньше всех сдает правильные ответы (или
наиболее правильные ответы), играет в основном
туре.
Для игры в основном туре команда выбирает из
своего состава ученика, который будет отвечать
на вопросы ведущего. Если у команды возникают
затруднения, окончательное решение принимает
либо ведущий, либо учитель.
В основном туре игроку нужно ответить на 10
вопросов, из которых первые 5 более простые, чем
остальные. Вопросы включают в себя основные
понятия, термины, факты из пройденного материала.
Ответив на 5 вопросов, ученик получает первое
«несгораемое» звание «бакалавр» науки,
определенной темой игры. В случае общего
проигрыша это звание остается за игроком (в
оценочном варианте ставится оценка «5»). Верные
ответы на следующие 5 вопросов тура приносят
игроку звание «магистр» с вручением диплома или
«медали» (в оценочном варианте – две отметки «5»).
У каждого игрока есть возможность использовать
три подсказки:
– 50 на 50: из четырех вариантов
ответов убираются 2 неверных;
– помощь команды: игрок подходит к своей команде
и в течение 30 с обсуждает с ней заданный
вопрос;
– помощь зала: ученики класса поднимают табличку
с той буквой, под которой, на их взгляд, находится
правильный ответ. (Помощники считают
количественное соотношение букв и объявляют
результат. Учитель может поощрить наиболее
результативного ученика, для чего помощники
фиксируют варианты ответов зала в списке
учителя.)
Продолжительность игры может составлять от 40 до 80 мин (от 3 до 6 отборочных и основных туров, соответственно). В конце игры подводятся итоги, награждаются победители.
«Кто станет магистром генетики?»
I отборочный тур
Расположите диплоидные хромосомные наборы животных в порядке их численного возрастания:
а) дрозофила;
б) черепаха болотная;
в) шимпанзе;
г) аскарида.

Ответ:
г) 2 хромосомы;
а) 8 хромосом;
в) 48 хромосом;
б) 50 хромосом.
Примечание . В этом и последующих отборочных турах участникам достаточно указать лишь последовательность букв, а данные (в этом случае – количество хромосом) приводить не надо.
I основной тур
1. Аутосомы – это:
а) дополнительные хромосомы в клетке;
б) клетки, способные воспроизводить себя;
в) органеллы, способные воспроизводить себя;
г) неполовые хромосомы.
2. Термин «ген» для обозначения наследственных задатков организма был предложен:
а) Иоганном Грегором Менделем;
б) Вильгельмом Людвигом Иогансеном;
в) Томасом Хантом Морганом;
г) Хуго Мари де Фризоле.
3. В клетках листьев сосны имеется 24 хромосомы; в эндосперме семян сосны их:
а) 12;
б) 24;
в) 36;
г) 48.
4. У всех млекопитающих, в том числе и у человека, гомогаметным является пол:
а) женский;
б) мужской;
в) мужской и женский;
г) в одних случаях – мужской, в других – женский.
5. У потомства могут появиться новые признаки, не свойственные родителям, т.к.:
а) при оплодотворении гаметы сливаются случайно;
б) в гаметах родителей часто происходят мутации;
в) у потомков родительские гены сочетаются в новых комбинациях;
г) одну половину генов потомство получает от матери, другую – от отца.
6. Аутбридинг – это:
а) изменение наследственных задатков при близкородственном скрещивании;
б) способ скрещивания, не приводящий к появлению новых признаков и свойств;
в) скрещивание особей одного вида, не состоящих в непосредственном родстве;
г) скрещивание особей разных видов.
7. У собак висячее ухо доминирует над стоячим. При скрещивании двух собак с висячими ушами получили 14 особей с висячими ушами и 4 со стоячими. Каковы генотипы скрещиваний:
а) АА х АА ;
б) Аа х АА ;
в) Аа х аа ;
г) Аа х Аа .
8. Голландский ученый Гуго де Фриз связал между собой теории двух ученых:
а) Дарвина и Ламарка;
б) Дарвина и Менделя;
в) Геккеля и Ламарка;
г) Геккеля и Менделя.
9. Какое потомство может быть получено от нормальной по признаку свертываемости крови гомозиготной женщины и мужчины-гемофилика:
а) мальчик – гемофилик, девочка – носитель дефектного гена;
б) здоровый по данному признаку мальчик, девочка – носитель дефектного гена;
в) здоровый мальчик, девочка – гемофилик;
г) здоровый мальчик, здоровая девочка.
10. После возникновения хромосомной мутации особь потеряла способность скрещиваться с особями своего вида. Следует ли считать эту особь новым видом:
а) да, т.к. главный критерий вида – репродуктивная изоляция;
б) да, т.к. хромосомная мутация может привести к коренному изменению характеристик данного организма;
в) нет, т.к. виды состоят из популяций, а эволюция идет при смене поколений; новый вид образуется тогда, когда из подобных особей сформируется популяция;
г) нет, т.к. хромосомная мутация не затрагивает генов и потому не играет роли в эволюции.
Ответы :
1 – г;
2 – б;
3 – а;
4 – а;
5 – в;
6 – в;
7 – г;
8 – б);
9 – б;
10 – в.
II отборочный тур
Расположите основные открытия в области генетики в хронологическом порядке:
а) закон относительной частоты встречаемости доминантных и рецессивных аллелей Харди–Вайнберга;
б) мутационная теория Гуго де Фриза;
в) законы наследования моно- и полигенных признаков Г.Менделя;
г) хромосомная теория наследования Т.Моргана.
Ответ :
в) 1855–1856 гг.;
б) 1901–1903 гг.;
а) 1908 г.;
г) 1910 г.
II основной тур
1. Ген находится в составе структуры:
а) РНК;
б) ДНК;
в) АТФ;
г) белка.
2. Основной метод исследования закономерностей наследственности и изменчивости, примененный Г.Менделем:
а) цитогенетический;
б) генеалогический;
в) статистический;
г) гибридологический.
3. Сцепленное наследование генов, локализованных в одной паре гомологичных хромосом, установил:
а) Г.Мендель;
б) Т.Морган;
в) В.Иогансен;
г) Г. де Фриз.
4. Трутни имеют хромосомный набор:
а) гаплоидный;
б) диплоидный;
в) триплоидный;
г) тетраплоидный.
5. Селекция в чистых линиях не имеет смысла, поскольку у всех индивидуумов:
а) одинаковый фенотип;
б) одинаковый генотип;
в) различный фенотип;
г) различный генотип.
6. Известно, что ген синдактилии (сращения указательного и среднего пальцев) локализован в Y-хромосоме. Как будет наследоваться эта аномалия у детей, родившихся от брака здоровой по этому признаку женщины и мужчины со сросшимися пальцами:
а) все дети будут с аномалией;
б) все дети будут здоровы, т.к. мать здорова по этому признаку;
в) девочки – здоровы, но каждый второй мальчик – с аномалией;
г) все мальчики будут с аномалией.
7. У мальчика IV группа крови, а у его сестры – I группа. О группах крови их родителей можно сказать:
а) оба родителя имеют IV группу крови;
б) один из родителей имеет I, а второй – IV группу крови;
в) один из родителей имеет II, другой – III группу крови;
г) у этих детей разные отцы.
8. У мальчика синдром Дауна. Какой была комбинация гамет при оплодотворении? Набор хромосом в гаметах:
1) (23 + Х);
2) (21 + Y);
3) (22 + ХХ);
4) (22 + Y).
Выберите правильный ответ:
а) 1 и 2;
б) 1 и 3;
в) 1 и 4;
г) 3 и 4.
9. Ген широколистности у некоторых растений является геном неполного доминирования по отношению к гену узколистности. При скрещивании широколистного и узколистного растений можно ожидать результат:
а) все широколистные;
б) все узколистные;
в) соотношение широколистных и узколистных 1:1;
г) все растения с листьями средней ширины.
10. Если бы в состав ДНК входило не 4, а 5 оснований, и генетический код был бы не триплетным, а тетраплетным (т.е. кодон состоял бы из четырех оснований), то общее возможное число кодонов составило бы:
а) 125;
б) 256;
в) 625;
г) 1024.
Ответы :
1 – б;
2 – г;
3 – б;
4 – а;
5 – б;
6 – г;
7 – в;
8 – в;
9 – г;
10 – в.
III отборочный тур
В каком порядке были присуждены Нобелевские премии за выдающиеся достижения и открытия фундаментальных законов генетики?
а) Роберту Холли, Хару Гобинду Коране и Маршаллу Ниренбергу за расшифровку генетического кода и его функций в синтезе белков;
б) Герману Дж.Мёллеру за открытие возникновения мутаций под воздействием рентгеновских лучей;
в) Томасу Ханту Моргану за открытие функций хромосом как носителей наследственности;
г) Фрэнсису Крику, Джеймсу Уотсону и Морису Уилкинсу за установление молекулярной структуры нуклеиновой кислоты и ее роли в передаче информации в живой материи.
Ответ :
в) 1933 г.;
б) 1946 г.;
г) 1962 г.;
а) 1968 г.
III основной тур
1. Число комбинаций гамет дигетерозиготных родительских особей при дигибридном скрещивании составляет:
а) 4;
б) 6;
в) 8;
г) 16.
2. Метод, не приемлемый в исследованиях генетики человека:
а) цитогенетический;
б) биохимический;
в) гибридологический;
г) генеалогический.
3. Закон, позволяющий определить частоты встречаемости доминантных и рецессивных генов в популяции по частоте встречаемости фенотипа, называется законом:
а) Г.Менделя;
б) Т.Моргана;
в) Н.Вавилова;
г) Харди–Вайнберга.
4. Если в ядрах клеток эпителия полости рта женщины обнаружено по одному Y-хроматину и не обнаружен X-хроматин, то ее хромосомный набор:
а) 45, Х0 ;
б) 46, ХХ ;
в) 46, XY ;
г) 47, XXY .
5. Совокупность всех наследственных задатков клетки или организма – это:
а) гены;
б) геном;
в) генотип;
г) генофонд.
6. Вид взаимодействия генов, при котором один ген не дает проявиться в фенотипе другой паре генов, называется:
а) комплементарностью;
б) плейотропией;
в) эпистазом;
г) полимерией.
7. Синдром Дауна вызывается появлением лишней хромосомы в 21-й паре. Это следует считать:
а) хромосомной мутацией;
б) генной мутацией;
в) геномной мутацией;
г) соматической мутацией.
8. Первая пара возможных родителей имеет группы крови II и III, вторая – IV и III. Ребенок имеет I группу крови. Кто является родителями ребенка:
а) может быть и I, и II пара;
б) не может быть ни I, ни II пара;
в) только I пара;
г) только II пара.
9. А – карий цвет глаз, а – голубой цвет глаз, В – темные волосы, в – светлые волосы. Вероятность рождения голубоглазого светловолосого ребенка у дигетерозиготных по этим признакам родителей равна:
а) 9/16;
б) 3/16;
в) 1/16;
г) 0.
10. Какое соотношение частоты генотипов АА , Аа и аа отвечает закону Харди–Вайнберга:
а) 0,25; 0,50; 0,25;
б) 0,36; 0,55; 0,09;
в) 0,64; 0,27; 0,09;
г) 0,29; 0,42; 0,29.
Ответы :
1.проявление ненаследственной изменчивости у особи зависит от ее генотипа поэтому пределы данной изменчивости ограничены 1)условиями среды 2)нормой1. – г);
2. – в);
3. – г);
4. – в);
5. – в);
6. – г);
7. – в);
8. – в);
9. – в);
10. – а).
реакции 3)фенотипом особи 4)случайными мутациями
2. нарушение последовательности нуклеотидов в молекуле ДНК-результат 1)геномной мутации 2)генной мутации 3)хромосомной мутации 4)нормы реакции
3.одним из предложений мутационной теории Х. Де Фриза является утверждение
1)мутации возникают под действием условий среды 2)мутации определённы 3)мутации носят групповой характер 4)мутации ненаправлены
4. комбинативную изменчивость обусловливают
1)мутации
2\)новые комбинации генов в результате мейоза и оплодотворения
3)новые комбинации признаков под влиянием условий среды
4)мутагены
акой признак является доминантным? Каковы генотипы всех членов этой семьи? 3.Мужчина, страдающий альбинизмом, женится на здоровой женщине, отец которой страдал альбинизмом. Каких детей можно ожидать от этого брака, если учесть, что альбинизм наследуется у человека как аутосомный рецессивный признак? 4. Голубоглазый мужчина женился на кареглазой женщине, мать которой имела голубые глаза. От этого брака родилась голубоглазая дочь и кареглазый сын. Определить генотипы всех членов семьи. 5. Сибирский длинношерстный кот Васька скрещивался с соседской кошкой Муркой. В результате этого скрещивания родились 4 короткошерстных и 2 длинношерстных котенка. Известно, что у кошек короткая шерсть - доминантный признак. Определить генотипы Васьки, Мурки и всех котят. 6. У человека встречается такая аномалия, как альбинизм (отсутствие пигментации). Среди индейцев Панамы она довольно широко распространена, несмотря на то, что браки между альбиносами запрещены законом. Как это можно объяснить? 7.* Рыжеволосая девушка выходит замуж за мужчину с не рыжими волосами, гомозиготного по этому признаку. Какова вероятность рождения от этого брака ребенка с не рыжими волосами, если известно, что рыжие волосы - рецессивный признак? 8.Так называемый «белый локон» или седая прядь у человека наследуется как доминантный аутосомный признак. Какова вероятность рождения детей без локона, если родители имели «белый локон» и были гетерозиготными по этому признаку? 9. Какое потомство по генотипу и фенотипу получится от скрещивания серого чистопородного кролика с черной крольчихой, если черный цвет меха это рецессивный признак? рой коровы - пять нормальных телят, от третьей коровы - пять бесшерстных телят. Определить генотипы скрещиваемых животных и их потомков. 10.* В живом уголке жили морские свинки: самец с длинной шерстью и такая же самка. От их скрещивания в потомстве появились свинки с длинной и короткой шерстью. Какова вероятность появления короткошерстных свинок, если скрестить самцов с длинной шерстью из первого поколения с короткошерстными самками? Короткая шерсть - рецессивный признак. 11. При скрещивании красноплодных томатов (доминантный признак) в потомстве было получено 87 красноплодных и 29 желтоплодных растений. Сколько среди них было гетерозигот? 12.Определить вероятность рождения детей без веснушек на лице (рецессивный признак), если известно следующее: женщина, имеющая веснушки на лице, отец которой был без веснушек, вступила в брак с мужчиной, имеющим веснушки на лице. Известно, что мать мужчины была без веснушек, а отец - с веснушками. 13.* У кукурузы темная окраска зерен доминирует над светлой. От скрещивания между собой гибридов получено 380 растений: из них 96 имели белые зерна, а остальные - темные. Сколько всего растений были гомозиготными по генотипу и сколько - гетерозиготными? 14. У пшеницы ген карликовости.доминирует над геном нормального роста. Каковы генотипы исходных форм, если в потомстве получилось расщепление по этому признаку в соотношении 1:1? При каких генотипах и фенотипах родительских форм получится фенотипически однородное потомство? 15. Предположим, что синдром Вернера (преждевременное старение человека) наследуется по аутосомно-рецессивному типу. Каков прогноз появления этого синдрома у потомков, если известно следующее: жена здорова, но ее отец имел синдром Вернера, муж также здоров, но его мать состарилась очень рано.
Часть А (выберите один правильный ответ)1. Грегор Мендель при изучении наследственности использовал метод:
А) генеалогический; Б) цитогенетический; В) гибридологический; Г) биохимический.
2. Взаимоисключающие состояния признака определяются генами:
А) доминантными; Б) аллельными; В) рецессивными; Г) альтернативными.
3. Взаимодействие неаллельных генов, при котором в потомстве проявляется новое состояние признака:
А) эпистаз; Б) комплементарность; В) полимерия; Г) плейотропия.
4. Комплекс генов, передаваемый потомству при размножении:
А) генотип; Б) фенотип; В) генофонд; Г) кариотип.
5. Наследование групп крови у человека относят к:
А) плейотропии; Б) кодоминированию; В) дигибридному скрещиванию; Г) комплементарности.
6. Признак, наследуемый сцеплено с полом:
А) острота зрения; Б) свертываемость крови; В) цвет волос; Г) число пальцев.
7. Модификационная изменчивость связана с изменением:
А) генотипа; Б) генофонда; В) фенотипа; Г) генома.
8. Пол, образующий гаметы, различающиеся по половой хромосоме:
А) гомогаметный; Б) гетерозиготный; В) гомозиготный; Г) гетерогаметный.
9. Анализирующим называют скрещивание:
А) двух гетерозигот; Б) доминантной гомозиготы и гетерозиготы; В) рецессивной гомозиготы и гетерозиготы;
Г) двух рецессивных гомозигот.
10. Гены, определяющие развитие альтернативных состояний признака:
А) доминантные; Б) аллельные; В) рецессивные; Г) мутантные.
11. Отсутствие пигмента в покровах птиц, земноводных, рептилий и млекопитающих – одно из доказательств закона:
А) чистоты гамет; Б) сцепленного наследования генов; В) гомологических рядов наследст.изменчивости; Г)доминирования
12. Изменения, происходящие с организмом под влиянием факторов внешней и внутренней среды, - это проявление:
А) мутационной изменчивости; Б) модификационной изменчивости; В) цитоплазматической изменчивости; Г) комбинативной.
13. Различия в размерах листьев одного дерева – это пример изменчивости:
А) генотипической; Б) модификационной; В) мутационной; Г) комбинативной.
14В бесполом размножении мхов и папоротников участвуют:
А) споры; Б) сперматозоиды; В) яйцеклетки; Г) пыльца.
15. Принципиальные различия между половым и бесполым размножением заключается в том, что половое размножение:
А) происходит только у высших растений; Б) это приспособление к неблагоприятным условиям среды; В) обеспечивает комбинативную изменчивость организма; Г) обеспечивает генетическое постоянство вида.
16. Клетки гаструлы:
А) гаплоидны; Б) диплоидны; В) тетраплоидны; Г) триплоидны.
17. Из эктодермы у млекопитающих образуются:
А) волосы и ногти; Б) скелетные мышцы; В) легкие; Г) хрящи.
18. Чистой линией называется:
А) потомство, не дающее разнообразия по изучаемому признаку; Б) разнообразное потомство, полученное от скрещивания разных особей; В) пара родителей, отличающихся друг от друга одним признаком; Г) особи одного вида.
19. У кареглазого мужчины и голубоглазой женщины родились трое кареглазых девочек и один голубоглазый мальчик. Ген карих глаз доминирует. Каковы генотипы родителей?
А) отец- АА, мать- Аа; Б) отец-аа, мать -АА; В) отец – аа, мать - Аа; Г) отец – Аа, мать - аа.
20. Организм с генотипом ВВСс образует гаметы:
А) В, С и с; Б) ВВ и Сс; В) ВС и Вс; Г) ВВС и ВВс.
21. Значение кроссинговера заключается в:
А) независимом распределении генов по гаметам; Б) сохранении диплоидного набора хромосом; В) создании новых наследственных комбинаций; Г) поддержании постоянства генотипов организма.
Часть В (выберите три правильных ответа)
1. Признаки, характеризующие генотип:
А) совокупность генов организма; Б) совокупность внешних признаков организма; В) совокупность внутренних качеств организма;
Г) совокупность хромосом; Д) совокупность половых клеток; Е) весь наследственный материал клетки.
2. Основные характеристики наследственной изменчивости:
А) генотипическая; Б) фенотипическая; В) индивидуальный характер;
Г) групповой характер; Д) зависит от условий среды; Е) сохраняется в потомстве.
Генетическое доказательство перекреста хромосом стало возможным благодаря открытию ряда генетических явлений - мутаций, гетерозиготного состояния и сцепления генов.
Первым генетическим объектом, на котором было установлено явление кроссинговера, была дрозофила. Для нее впервые был составлен систематический указатель генов, находящихся в разных хромосомах, и выявлены все группы сцепления.
Рассмотрим один из классических опытов Т. Моргана по перекресту, позволивший ему доказать наследственную дискретность хромосом.
Первые опыты по кроссинговеру
При скрещивании мух, различающихся по двум парам признаков, серых с рудиментарными крыльями (vg - vestigial) b + vg/b + vg и черных (b - black) с нормальными крыльями b vg + /b vg + в F 1 дигетерозиготные особи b + vg/b vg + по фенотипу оказываются серыми с нормальными крыльями.
На рисунке представлены два направления скрещиваний: в одном дигетерозиготой является самец, в другом - самка. Если гибридные самцы скрещиваются с самками, рецессивными по обоим генам, т. е. производится анализирующее скрещивание b vg/b vg x b + vg/b vg + , то в потомстве получается расщепление в отношении 1 (b + vg/b vg) x 1 (b vg + x b vg).
Такое расщепление показывает, что данный дигибрид образует только два сорта гамет b + vg и bvg + вместо четырех, причем сочетание генов в гаметах соответствует тому, которое было у родителей. Исходя из указанного расщепления, следует предположить, что у самца не происходит обмена участками гомологичных хромосом. В дальнейшем выяснилось, что у самцов дрозофилы действительно как в аутосомах, так и в половых хромосомах и норме нет перекреста. Поэтому при описанном анализирующем скрещивании в потомстве восстанавливаются две исходные родительские комбинации признаков: мухи с черной окраской тела и нормальными крыльями и мухи с серой окраской тела и рудиментарными крыльями. При этом они оказываются в равных числовых отношениях независимо от пола. В данном случае мы имеем пример полного сцепления генов, находящихся в одной паре гомологичных аутосом.
В реципрокном скрещивании дигетерозиготных самок с самцом анализатором, гомозиготным по этим же двум рецессивным генам, в потомстве наблюдается иное расщепление. Кроме родительских комбинаций признаков, в потомстве появляются новые типы - мухи с черным телом и короткими (рудиментарными) крыльями, а также мухи с серым телом и нормальными крыльями. Следовательно, в этом скрещивании сцепление генов нарушается. Гены в гомологичных хромосомах поменялись местами благодаря перекресту хромосом.
Гаметы с хромосомами, претерпевшими кроссинговер, называют кроссоверными , а не претерпевшие такового - некроссоверными . Соответственно организмы, возникшие в потомстве анализирующего скрещивания от сочетания кроссоверных гамет с гаметами анализатора, называют кроссоверными , или рекомбинантными , а возникшие от сочетания некроссоверных гамет с гаметами анализатора - некроссоверными , или нерекомбинантными .
При анализе расщепления в случае кроссинговера обращает на себя внимание определенное числовое отношение особей разных классов. Обе исходные родительские комбинации признаков, образовавшиеся из некроссоверных гамет, оказываются в потомстве анализирующего скрещивания в равном числовом отношении. В указанном опыте с дрозофилой тех и других особей было примерно по 41,5%. В сумме некроссоверные особи составили 83% от общего числа особей в потомстве. Два кроссоверных класса по числу особей также одинаковы, и сумма их равна 17%.
В качестве другого классического примера сцепления и перекреста можно привести опыт К. Гетчинсона, проведенный на кукурузе в 20-х годах. Были скрещены две гомозиготные линии кукурузы, одна из которых имела зерна с окрашенным алейроном и гладким эндоспермом. Эти признаки определяются доминантными генами c + c + sh + sh + . Другая линия имела рецессивные аллели этих генов c c sh sh, которые определяли соответственно признаки: неокрашенный алейрон и морщинистый эндосперм. Эти пары аллелей находятся в одной паре гомологичных хромосом.
Скрещивание указанных линий между собой c + sh + /c + sh + x с sh/c sh дает гетерозиготу c + sh + /c sh.
Расщепление при анализирующем скрещивании c + sh + /с sh x c sh/c sh так же, как в опыте с дрозофилой, оказывается не соответствующим независимому поведению каждой пары аллелей. При этом расщеплении количество некроссоверных зерен в початке оказывается 96,4%, а кроссоверных - 3,6%.

Результаты опытов на дрозофиле и кукурузе показывают, что действительно существует сцепление генов, и лишь в известном проценте случаев оно нарушается вследствие кроссинговера. Отсюда и следует первое положение о перекресте хромосом, которое гласит, что между гомологичными хромосомами может осуществляться взаимный обмен идентичными участками. Гены, находящиеся в идентичных участках гомологичных хромосом, перемещаются из одной гомологичной хромосомы в другую.
Следовательно, в то время как независимое комбинирование генов, находящихся в негомологичных хромосомах, определяется случайным их расхождением в редукционном делении, рекомбинация сцепленных генов обеспечивается процессом перекреста гомологичных хромосом.
Величина перекреста и линейное расположение генов в хромосоме
Величина кроссинговера измеряется отношением числа кроссоверных особей к общему числу особей в потомстве анализируемого скрещивания и выражается в процентах.
Рекомбинация происходит реципрокно, т. е. между родительскими хромосомами осуществляется взаимный обмен; это и обязывает подсчитывать кроссоверные классы вместе как результат одного события.
Величина перекреста хромосом отражает силу сцепления генов в хромосоме: чем больше величина перекреста, тем меньше сила сцепления. Т. Морган предположил, что частота кроссинговера показывает относительное расстояние между генами: чем чаще осуществляется кроссинговер, тем далее отстоят гены друг от друга в хромосоме, чем реже кроссинговер, тем они ближе друг к другу.
Когда мы указываем, что рекомбинация генов черного цвета тела и коротких крыльев у дрозофилы происходит с частотой 17%, то эта величина определенным образом характеризует расстояние между данными генами в хромосоме. То же самое относится к случаю перекреста хромосом у кукурузы, где 3,6% рекомбинаций показывают частоту осуществленных обменов между двумя участками Гомологичных хромосом.
На основе многочисленных генетических исследований Морган выдвинул гипотезу линейного расположения генов в хромосоме. Только при этом допущении процент рекомбинантов может отражать относительное расстояние между генами в хромосоме.
Одним из классических генетических опытов Моргана, доказывающих линейное расположение генов, был следующий опыт с дрозофилой. Самки, гетерозиготные по трем сцепленным рецессивным генам, определяющим желтый цвет тела y (yellow), белый цвет глаз w (white) и вильчатые крылья bi (bifid), были скрещены с самцами, гомозиготными по этим трем генам. В потомстве было получено 1160 мух некроссоверных (нормальных и одновременно несущих все три рецессивных признака), 15 мух кроссоверных, возникающих от перекреста между генами у и w, и 43 особи от кроссинговера между генами w и bi. Полученные результаты в процентах перекреста между генами представляли следующее соотношение:

Из этих данных с очевидностью вытекает, что процент перекреста является функцией расстояния между генами и их последовательного, т. е. линейного, расположения в хромосоме. Расстояние между генами y и bi равно сумме двух одинарных перекрестов между y и w, w и bi.
Воспроизводимость этих результатов в повторных опытах указывает на то, что местоположение генов вдоль по длине хромосомы строго фиксировано, т. е. каждый ген занимает в хромосоме свое определенное место - локус.
Одинарный и множественный перекресты хромосом
Приняв положения, что 1) генов в хромосоме может быть много, 2) гены расположены в хромосоме в линейном порядке, 3) каждая аллельная пара занимает определенные и идентичные локусы в гомологичных хромосомах, Т. Морган допустил, что перекрест между гомологичными хромосомами может происходить одновременно в нескольких точках.
Это предположение было им доказано на дрозофиле, а затем полностью подтверждено на ряде животных, растительных объектов и микроорганизмов.
Перекрест, происходящий лишь в одном месте, называют одинарным перекрестом, в двух точках одновременно - двойным, в трех - тройным и т. д., т. е. кроссинговер может быть множественным. Пусть, например, в гомологичной паре хромосом содержатся три пары аллелей в гетерозиготном состоянии: ABC/abc.
Тогда перекрест, произошедший только в участке между генами А и В или между В и С (в разных клетках), будет одинарным. В результате одинарного перекреста возникают в каждом случае только две кроссоверные хромосомы (гаметы), а именно аВС и Abc или АВс и аbС.
Если каждая из этих хромосом будет сочетаться в зиготе с гомологичной хромосомой, несущей все три рецессивные аллели а, b и с, то в потомстве получатся следующие генотипы кроссоверных зигот: аВС/abc и Abc/abc или АВс/abc и аbС/abc.
По проценту кроссоверных классов устанавливают частоту произошедших одинарных обменов между генами А и В или В и С.
В результате одновременного перекреста между А и В и между В и С происходит обмен средним участком хромосомы - двойной обмен. При этом в гетерозиготе возникает новый сорт гамет с кроссоверными хромосомами АbС и аВс, которые выявляются с помощью анализирующего скрещивания. В потомстве появляются зиготы со следующей комбинацией генов: AbC/abc и aBc/abc.
Одинарный и двойной перекресты между гомологичными хромосомами доказываются следующим генетическим анализом. В таблице рассмотрен конкретный опыт на дрозофиле, в котором общее число кроссоверных и некроссоверных особей составляло 521. Этот анализ представлен в общей форме, без указания определенных генов, с тем, чтобы подчеркнуть его принципиальное значение.
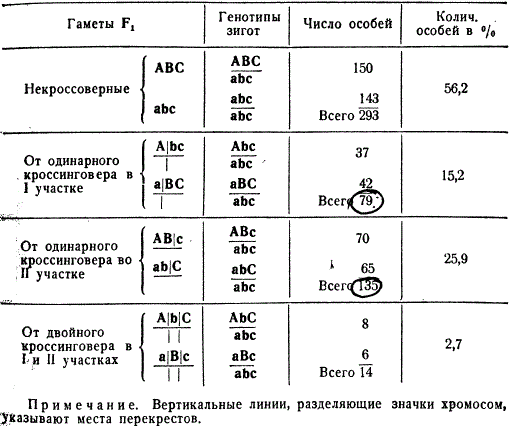
Чтобы вычислить процент одинарного перекреста в обоих участках необходимо к одинарным кроссоверам 79 и 135 прибавить число дробей, полученных при двойном перекресте, так как последний произошел как в первом, так и во втором участке.
Рассчитаем процент перекреста между генами A и B: 79 + 14 = 93, 93:521⋅100 = 17,9%.
Один процент перекреста был принят за единицу измерения перекреста, в русской литературе он получил название морганиды.
Следуя тому же методу подсчета перекреста для второго участка - между генами В и С, получим 28,6%, или морганиды. Таким образом, мы определили относительные расстояния между генами: расстояние между А и В - 17,9 и между В и С - 28,6 единиц перекреста, т. е. морганид.
Если правильно, что перекрест есть функция расстояния между генами, то нам легко установить расстояние между генами А и С, так как оно должно быть примерно равно сумме двух частот одинарного перекреста: 17,9 + 28,6 = 46,5. Однако общее число одинарных кроссоверов между генами А и С составляет 214 (79 + 135) особей, или 41,1 морганид, т. е. расстояние между генами А и С, рассчитанное нами ранее, оказалось больше на 46,5 - 41,1 = 5,4 морганиды. Такое расхождение как будто противоречит предыдущему опыту с генами y w bi, где частота перекреста (4,7%) между крайними генами (y и bi) точно совпала с суммой частот перекреста между генами y и w (1,2%) и w и bi (3,5%). Но в том случае гены находятся на близком расстоянии друг от друга, а в примере с генами ABC гены расположены на большом расстоянии друг от друга.
Расхождение в подсчетах объясняется тем, что между далеко отстоящими генами может происходить двойной перекрест, который затрудняет оценку истинного расстояния между генами. Двойной кроссинговер можно не заметить, если расстояние между генами А и С не маркировано третьим геном В.
В случае произошедшего двойного обмена участками внутри хромосом, например, гены А и С останутся на своих местах, и обмен между ними не будет обнаружен. При этом, чем дальше отстоят друг от друга в хромосоме гены А и С, тем больше вероятность двойных перекрестов между ними. Процент рекомбинаций между двумя генами тем точнее отражает расстояние между ними, чем оно меньше, так как в случае малого расстояния уменьшается возможность двойных обменов. Поэтому кроссинговер между генами А и С (41,1%) без учета двойных кроссоверов оказывается меньше, чем сумма единиц кроссинговера между генами А и В, а также между В и С (46,5%).
Для учета двойного кроссинговера необходимо иметь дополнительный метчик, находящийся между двумя изучаемыми генами. В рассмотренном примере таким метчиком является ген В. Определение расстояния от А до С осуществляют следующим образом: к сумме процентов одинарных кроссоверных классов (41,1%) прибавляют удвоенный процент двойных кроссоверов (2,7 X 2 = 5,4%). Удвоение процента двойных кроссоверов необходимо в связи с тем, что каждый двойной кроссинговер возникает благодаря двум независимым одинарным разрывам в двух точках. Чтобы вычислить процент одинарного кроссинговера, необходимо величину двойного кроссинговера умножить на 2. В рассматриваемом примере в итоге получается 41,1 + 5,4 = 46,5%, что равняется сумме, полученной от сложения процентов кроссинговера в двух участках: от А до В и от В до С.
Расчет процента кроссинговера между двумя генами можно производить не только на основе данных анализирующего скрещивания, но и по результатам расщепления в F 2 . Для простоты объяснения предположим, что нам известен процент перекреста между генами А и В и что он равен 20%. Тогда в F 1 дигетерозигота AB/ab должна образовать гаметы в следующих соотношениях: 0,4АВ: 0,1Аb: 0,1аВ: 0,4аb (так как кроссоверных гамет 20%, а некроссоверных 80%). В F 2 особи, гомозиготные по обоим рецессивным генам, возникают только в результате слияния двух гамет ab с частотой 0,4 X 0,4 = 0,16. В любом случае процент гамет с двумя рецессивными генами у особей F 1 определяется как корень квадратный из частоты класса ab в F 2 , выраженной в долях морганид. В том случае, когда производится скрещивание типа AB/AB x ab/ab, определенная по F 2 частота гамет ab, образуемых дигетерозиготой F 1 равна половине частоты всех некроссоверных гамет. Если же осуществляется скрещивание типа Ab/Ab x aB/aB, то определенная по F 2 частота гамет ab, образуемых гибридом F 1 равна половине частоты всех кроссоверных гамет.
Мейоз и оплодотворение обеспечивают получение организмами нового поколения эволюционно сложившегося, сбалансированного по дозам генов наследственного материала, на основе которого осуществляется развитие организма и отдельных его клеток. Благодаря этим двум механизмам в ряду поколений особей данного вида формируются определенные видовые характеристики и вид как реальная единица живой природы существует продолжительное время. Однако у разных представителей вида в силу постоянно идущего мутационного процесса один и тот же набор генов генома представлен разными их аллелями. Так как при половом размножении у многих видов в воспроизведении потомства принимают участие две особи, то совершенно очевидно, что в результате оплодотворения разные зиготы получают неодинаковый набор аллелей в их генотипах. Увеличению генотипического разнообразия представителей вида способствуют также механизмы, приводящие к перекомбинации родительских аллелей особи в ее гаметах. Действительно, если бы гаметы, образуемые организмом, были одинаковы по набору аллелей в их геноме, то у потомков одной пары организмов при раздельнополости или одного гермафродитного организма не наблюдалось бы генотипического разнообразия. В каждом новом поколении вида генотипически различными были бы лишь дети разных родителей.
Реально в природе наблюдается разнообразие потомков одних и тех же родителей. Например, родные братья и сестры различаются не только по полу, но и по другим признакам. Такие различия потомков объясняются тем, что в каждом акте оплодотворения встречаются генетически различающиеся гаметы. Механизмом,обеспечивающим разнообразие гамет, образуемых одним и тем же организмом, является мейоз, в ходе которого происходит не только уменьшение вдвое наследственного материала, попадающего в гаметы, но и эффективное перераспределение родительских аллелей между гаметами. Процессами, приводящими к перекомбинации генов и целых хромосом в половых клетках, являются Кроссинговер и расхождение бивалентов в анафазе I мейоза (см. гл. 5).
Кроссинговер. Этот процесс происходит в профазе I мейоза в то время, когда гомологичные хромосомы тесно сближены в результате конъюгации и образуют биваленты. В ходе кроссинговера осуществляется обмен соответствующими участками между взаимно переплетающимися хроматидами гомологичных хромосом (рис. 3.72). Этот процесс обеспечивает перекомбинацию отцовских и материнских аллелей генов в каждой группе сцепления. В разных предшественниках гамет Кроссинговер происходит в различных участках хромосом, в результате чего образуется большое разнообразие сочетаний родительских аллелей в хромосомах.
Рис. 3.72. Кроссинговер как источник генетического разнообразия гамет:
I - оплодотворение родительских гамет а и б с образованием зиготы в; II - гаметогенез в организме, развившемся из зиготы в ; г - кроссинговер, происходящий между гомологами в профазе I; д - клетки, образовавшиеся после 1-го мейотического деления; е, ж - клетки, образовавшиеся после 2-го деления мейоза (е - некроссоверные гаметы с исходными родительскими хромосомами; ж - кроссоверные гаметы с перекомбинацией наследственного материала в гомологичных хромосомах)
Понятно, что кроссинговер как механизм рекомбинации эффективен лишь в том случае, когда соответствующие гены отцовской и материнской хромосом представлены разными аллелями. Абсолютно идентичные группы сцепления при кроссинговере не дают новых сочетаний аллелей.
Кроссинговер происходит не только в предшественницах половых клеток при мейозе. Он наблюдается также в соматических клетках при митозе. Соматический кроссинговер описан у дрозофилы, у некоторых видов плесеней. Он осуществляется в ходе митоза между гомологичными хромосомами, однако его частота в 10 000 раз меньше частоты мейотического кроссинговера, от механизма которого он ничем не отличается. В результате митотического кроссинговера появляются клоны соматических клеток, различающихся по содержанию в них аллелей отдельных генов. Если в генотипе зиготы данный ген представлен двумя разными аллелями, то в результате соматического кроссинговера могут появиться клетки с одинаковыми либо отцовскими, либо материнскими аллелями данного гена (рис. 3.73).

Рис. 3.73. Кроссинговер в соматических клетках:
1 - соматическая клетка, в гомологичных хромосомах которой ген А представлен двумя разными аллелями (А и а); 2 - кроссинговер; 3 - результат обмена соответствующими участками между гомологичяыми хромосомами; 4 - расположение гомологов в плоскости экватора веретена деления в метафазе митоза (два варианта); 5 - образование дочерних клеток; 6 - образование гетерозитотиых по гену А клеток, сходных с материнской клеткой по набору аллелей (Аа); 7 - образование гомозиготных по гену А клеток, отличающихся от материнской клетки по набору аллелей (АА или аа)
Расхождение бивалентов в анафазе I мейоза. В метафазе I мейоза в экваториальной плоскости ахромативнового веретена выстраиваются биваленты, состоящие из одной отцовской и одной материнской хромосомы. Расхождение гомологов, которые несут разный набор аллелей генов в анафазе I мейоза, приводит к образованию гамет, отличающихся по аллельному составу отдельных групп сцепления (рис. 3.74).

Рис. 3.74. Расхождение гомологичных хромосом в анафазе I мейоза
как источник генетического разнообразия гамет:
1 -метафаза I мейоза (расположение бивалента в плоскости экватора веретена деления); 2 - анафаза I мейоза (расхождение гомологов, несущих разные аллели гена А к разным полюсам); 3 - второе мейотическое деление (образование двух типов гамет, различающихся по аллелям гена А)

Рис. 3.75. Случайный характер расположения бивалентов в метафазе (1 )
и независимое расхождение их в анафазе (2 ) первого мейотического деления
В связи с тем что ориентация бивалентов по отношению к полюсам веретена в метафазе I оказывается случайной, в анафазе I мейоза в каждом отдельном случае к разным полюсам направляется гаплоидный набор хромосом, содержащий оригинальную комбинацию родительских групп сцепления (рис. 3.75). Разнообразие гамет, обусловленное независимым поведением бивалентов, тем больше, чем больше групп сцепления в геноме данного вида. Оно может быть выражено формулой 2 n , где п - число хромосом в гаплоидном наборе. Так, у дрозофилы п = 4 и количество типов гамет, обеспечиваемое перекомбинацией родительских хромосом в них, равно 2 4 = 16. У человека п = 23, и разнообразие гамет, обусловленное этим механизмом, соответствует 2 23 , или 8388608.
Кроссинговер и процесс расхождения бивалентов в анафазе I мейоза обеспечивают эффективную рекомбинацию аллелей и групп сцепления генов в гаметах, образуемых одним организмом.
Оплодотворение. Случайная встреча разных гамет при оплодотворении приводит к тому, что среди особей вида практически невозможно появление двух генотипически одинаковых организмов. Достигаемое с помощью описанных процессов генотипическое разнообразие особей предполагает наследственные различия между ними на базе общего видового генома.
Таким образом, геном как высший уровень организации наследственного материала благодаря мейозу и оплодотворению сохраняет свои видовые характеристики. Но одновременно эти же процессы обеспечивают индивидуальные наследственные различия особей, в основе которых лежит рекомбинация генов и хромосом, т.е. комбинативную изменчивость. Комбинативная изменчивость, проявляющаяся в генотипическом разнообразии особей, повышает выживаемость вида в изменяющихся условиях его существования.
В 1909 г. бельгийский цитолог Янссенс наблюдал образование хиазм во время профазы I мейоза. Генетическое значение этого процесса разъяснил Морган, высказавший мнение, что кроссинговер (обмен аллелями) происходит в результате разрыва и рекомбинации гомологичных хромосом во время образования хиазм. В это время части двух хромосом могут перекрещиваться и обмениваться своими участками. В результате возникают качественно новые хромосомы, содержащие участки (гены) как материнских, так и отцовских хромосом. Аллели, входящие в группы сцепления у родительских особей, разделяются и образуются новые сочетания, которые попадают в гаметы, - процесс, называемы генетической рекомбинацией. Потомков, которые получаются из таких гамет с "новыми" сочетаниями аллелей, называют рекомбинантными.
Частота (процент) перекреста между двумя генами, расположенными в одной хромосоме, пропорциональна расстоянию между ними. Кроссинговер между двумя генами происходит тем реже, чем ближе друг к другу они расположены. По мере увеличения расстояния между генами все более возрастает вероятность того, что кроссинговер разведет их по двум разным гомологичным хромосомам.
Гибриды первого поколения (самки) были скрещены с чернотелыми зачаточнокрылыми самцами. В F2 кроме родительских комбинаций признаков, появились новые - мухи с черным телом и зачаточными крыльями, а также с серым телом и нормальными крыльями. Правда, количество рекомбинантных потомков невелико и составляет 17%, а родительских - 83%. Причиной появления небольшого количества мух с новыми сочетаниями признаков является кроссинговер, который приводит к новому рекомбинантному сочетанию аллелей генов b+ и vg в гомологичных хромосомах. Эти обмены происходят с вероятностью 17% и в итоге дают два класса рекомбинантов с равной вероятностью - по 8,5%.
Биологическое значение кроссинговера чрезвычайно велико, поскольку генетическая рекомбинация позволяет создавать новые, ранее не существовавшие комбинации генов и тем самым повышать наследственную изменчивость, которая дает широкие возможности адаптации организма в различных условиях среды.
Мутагены и их тестирование
Мутагены - физические и химические факторы воздействие которых на живые
организмы вызывает изменения наследственных свойств (генотипа). Мутагены
разделяются на: физические (рентгеновские и гамма-лучи. радионуклиды,
протоны, нейтроны и пр.), физико-химические (волокна, асбест), химические
(пестициды, минеральные удобрения, тяжелые металлы и др.). биологические
(некоторые вирусы, бактерии).
Тестирование на мутагенность. Стратегия тестирования на мутагенность. Для тестирования всех веществ, с которыми на протяжении жизни человек может контактировать, потребовался бы непомерно большой объем работы, поэтому была признана необходимость первоочередной проверки на мутагенность лекарств, пищевых добавок, пестицидов, гербицидов, инсектицидов, косметических средств, наиболее широко распространенных загрязнителей воды и воздуха, а также производственных вредностей. Второй методологический принцип заключается в выборочности тестирования. Это означает, что вещество анализируется на мутагенность при наличии двух обязательных условий: распространенности в среде обитания человека и наличии структурного сходства с известными мутагенами или канцерогенами. Отсутствие универсального теста, позволяющего одномоментно регистрировать индукцию изучаемым веществом (и его возможными метаболитами) различных категорий мутаций в половых и соматических клетках, служит основанием третьего принципа - комплексного использования специализированных тест-систем. Наконец, четвертый методологический принцип подразумевает ступенчатость тестирования веществ на мутагенную активность. Этот принцип берет начало от одной из первых и наиболее известных схем, предложенной в 1973 г. Б, Бриджесом и предусматривавшей три последовательных этапа исследования. 1. На первом этапе мутагенные свойства вещества изучали простыми и быстро выполнимыми методами (с использованием микроорганизмов и дрозофилы в качестве тест-объектов) для определения его способности индуцировать генные мутации. Выявление такой способности предполагало запрет на применение данного вещества. 2. В случае особой медицинской или экономической значимости мутагена его тестировали на млекопитающих in vivo. Аналогичное исследование проводилось также для веществ, не продемонстрировавших мутагенных свойств в тестах первого этапа. Если исследуемый агент не проявлял мутагенных свойств, постулировалась безопасность применения его человеком. Вещества, проявившие мутагенность, либо запрещали для использования, либо, если они относились к категории особо значимых, или незаменимых, исследовали дополнительно. 3. На заключительном этапе проводили тестирование для установления количественных закономерностей мутагенного действия таких специфических веществ и оценку риска применения их человеком. Данная схема послужила прототипом целого ряда методик комплексного тестирования на мутагенность. Принципиально новым шагом на пути развития этого направления следует считать программу, предложенную в 1996 г. Дж. Эшби с соавторами, Исключительно важной особенностью этой программы является ее направленность не только на оценку мутагенности тестируемого вещества, но и на прогноз канцерогенности данного химического соединения и возможного механизма канцерогенеза. Современная система доказательств взаимосвязи между процессами мутагенеза и канцерогенеза включает целый ряд экспериментальных подтверждений обсуждаемой проблемы. Среди них: 1) наличие хорошо изученных наследственных заболеваний, при которых одновременно с повышенной чувствительностью к действию мутагенов наблюдается многократное превышение средней частоты возникновения злокачественных новообразований; 2) четко установленная сопряженность мутагенного и канцерогенного действия противоопухолевых цитостатиков, индуцирующих мутации в соматических клетках и за счет этого оказывающих терапевтическое воздействие, но способных вызывать у леченных онкологических больных развитие вторичных опухолей; 3) накопленные сведения о возможной активации протоонкогенов за счет индукции генных и хромосомных мутаций; 4) описание случаев спорадических моногенных доминантных мутаций, обусловливающих развитие опухолей различных органов. В программе Дж. Эшби постулируется, что вещество не является канцерогеном, если оно не проявляет мутагенного и генотоксического действия in vivo. Те же вещества, которым названные эффекты свойственны, являются потенциальными генотоксическими канцерогенами.
Билет 6
Цитологические доказательства опытов Штерна. Рисовать.
Опыт Штерна.К Х-хромосоме был добавлен фрагмент У-хромосомы, и она получила Г-образную форму. В начале 30-х годов К. Штерн получил линии дрозофилы, имеющие половые хромосомы, отличимые друг от друга на цитологическом уровне. У самки на одну из Х-хромосом был перенесен небольшой фрагментY- хромосомы, что придало ей специфическую Г-образную форму, легко различимую под микроскопом
Схема опыта по цитологическому доказательству кроссинговера на D. melanogaster
Были получены самки, гетерозиготные по двум указанным морфологически различным X-хромосомам и одновременно по двум генам Ваг (В) и carnation {car).
Цитологический анализ 374 препаратов самок показал, что в 369 случаях кариотип соответствовал ожидаемому. Все четыре класса самок имели по одной нормальной, т.е. палочковидной Х-хромосоме, полученной от отца. Кроссоверные (т.е. В саг + по фенотипу) самки содержали двуплечую Г-образную Х-хромосому.
Определение летальных рецессивных мутаций (метод CBL и Меллер 5)
Летальные гены - вызывающие смертельный исход в гомозиготном состоянии. Наряду с ними известно большое число полулетальных факторов, приводящих очень часто к рождению различного рода нежизнеспособных уродов или просто тем или иным способом отражающихся на жизнеспособности организмов. В наст, время Л. г. известны у дрозофилы, мышей, кроликов, собак, свиней, овец, лошадей, рогатого скота, птиц, у ряда растений, у человека и т. д. Примером полулетального фактора у человека может служить гемофилия, при наличии которой вместо нормального свертывания крови в 5-5х/2 мин. этот процесс иногда затягивается до 120 мин. и даже более; летальный ген гемофилии локализован в половой хромосоме, чем и объясняется передача этого признака половине сыновей от внешне здоровой матери, являющейся гетерозиготной по этому фактору
Наиболее удобные методы учета мутаций разработаны для дрозофилы. Собственно именно создание методов учета рецессивных летальных мутаций в Х-хромосоме определило успех Г.Меллера, открывшего действие рентгеновых лучей на мутационный процесс у дрозофилы. Для учета рецессивных летальных мутаций, сцепленных с полом, у дрозофилы широко применяют метод Меллер-5. Самки линии Меллер-5, или М-5, несут в обеих Х-хромосомах по две инверсии: sc 8 и сигма49. Инверсия sc 8 захватывает почти всю Х-хромосому, а в ее пределах находится еще одна инверсия –сигма49. В этой системе кроссинговер полностью подавлен. Используемые инверсии не имеют рецессивного летального действия. Кроме того, обе хромосомы М-5 несут три маркера: два рецессивных – w a (абрикосовый цвет глаз) и sc 8 (укороченные щетинки-фенотипическое проявление одноименной инверсии, затрагивающий ген sc) и один доминантный – Bar. При скрещивании исследуемых самцов с самками М-5 в индивидуальных семьях F 2 получают по два класса самок и самцов, если в Х-хромосоме сперматозоидов исходного самца не возникла рецессивная летальная мутация. Если же рецессивная леталь появилась, то в соответствующей индивидуальной культуре в F 2 мы получим только один класс самцов, будут отсутствовать самцы дикого типа w + B + . Метод Меллер-5 можно использовать и для регистрации рецессивных мутаций в Х-хромосоме с видимым проявлением. Для этой цели удобнее применять метод Double yellow, основанный на скрещивании исследуемых самцов с самками, несущими сцепленные Х-хромосомы. Благодаря тому, что при таком скрещивании сыновья получают свою Х-хромосому непосредственно от отца, рецессивные мутации в этой хромосоме можно учитывать уже в F 1 . Учет летальных мутаций и мутаций с видимым фенотипическим проявлением легче удается для Х-хромосом дрозофилы благодаря специфике ее наследования. Однако существуют методы учета летальных мутаций в аутосомах. Например, для учета рецессивных летальных мутаций в хромосоме 2 используют так называемый метод сбалансированных леталей. Для этого применяют линию, гетерозиготную по хромосоме 2. В одном гомологе находятся доминантные гены Cyrly (Cy-загнутые крылья) и Lobe (L- уменьшение глаза лопастной формы), в другом гомологе Plum (Pm- сливово-коричневый цвет глаз). Кроме того, хромосому Су L содержит инверсии, припятствующие кроссинговеру. Все три доминантные мутации обладают рецессивным летальным действием. Благодаря этому при разведении такой линии выживают только гетерозиготы по указанным генам. Это и есть система сбалансированных леталей. Для изучения рецессивных летальных мутаций, а также рецессивных мутаций с видимым проявлением исследуемых мух скрещивают с мухами CyL/Pm. В F 1 получают мух, гетерозиготных по той или другой хромосоме исследуемой линии, и индивидуально вновь скрещивают сегрегантов CyL с мухами CyL/Pm. В F 2 скрещивают между собой самцов и самок с признаками CyL и анализируют F 3 . В отсутствие рецессивной летальной мутации расщепление F 3 будет 2CyL:1Cy + L + , а если в половых клетках мух исходной линии возникали летальные мутации, то в соответствующих индивидуальных культурах в F 3 не будет нормальных мух 2CyL:0Cy + L + . Аналогично учитывают в F 3 и рецессивные мутации с видимым проявлением в хромосоме 2.
Билет 7
Комбинативная изменчивость и ее значение.
Комбинативной называют изменчивость, в основе которой лежит образование рекомбинаций, т.е. таких комбинаций генов, которых не было у родителей.
В основе комбинативной изменчивости лежит половое размножение организмов, вследствие которого возникает огромное разнообразие генотипов. Практически неограниченными источниками генетической изменчивости служат три процесса:
Независимое расхождение гомологичных хромосом в первом мейотическом делении. Именно независимое комбинирование хромосом при мейозе является основой третьего закона Менделя. Появление зеленых гладких и желтых морщинистых семян гороха во втором поколении от скрещивания растений с желтыми гладкими и зелеными морщинистыми семенами – пример комбинативной изменчивости.
Взаимный обмен участками гомологичных хромосом, или кроссинговер. Он создает новые группы сцепления, т.е. служит важным источником генетической рекомбинации аллелей. Рекомбинантные хромосомы, оказавшись в зиготе, способствуют появлению признаков, нетипичных для каждого из родителей.
Случайное сочетание гамет при оплодотворении.
Эти источники комбинативной изменчивости действуют независимо и одновременно, обеспечивая при этом постоянную «перетасовку» генов, что приводит к появлению организмов с другими генотипом и фенотипом (сами гены при этом не изменяются). Однако новые комбинации генов довольно легко распадаются при передаче из поколения в поколение.
Источники :
Кроссинговер при мейозе (гомологичные хромосомы тесно сближаются и меняются участками). Кроссинговер происходит в начале мейоза, когда гомологичные хромосомы выстраиваются друг против друга. При этом участки гомологичных хромосом перекрещиваются, отрываются, а затем вновь присоединяются, но уже к другой хромосоме. В конечном итоге образуются четыре хромосомы с разными комбинациями генов. Хромосомы, называемые «рекомбинантными», несут новые комбинации генов (Ab и аВ), отсутствовавшие в исходных хромосомах (АВ и ab) - Независимое расхождение хромосом при мейозе (каждая пара гомологичных хромосом расходится независимо от других пар). - Случайное слияние гамет при оплодотворении.
Комбинативная изменчивость является важнейшим источником всего колоссального наследственного разнообразия, характерного для живых организмов. Однако перечисленные источники изменчивости не порождают существенных для выживания стабильных изменений в генотипе, которые необходимы, согласно эволюционной теории, для возникновения новых видов. Такие изменения возникают в результате мутаций.
Комбинативная изменчивость объясняет, почему у детей обнаруживаются новые сочетания признаков родственников по материнской и отцовской линиям, причём в таких конкретных вариантах, которые не были свойственны ни отцу, ни матери, ни дедушке, ни бабушке и т.д. Благодаря комбинативной изменчивости создаётся разнообразие генотипов в потомстве, что имеет большое значение для эволюционного процесса в связи с тем, что: 1) увеличивается разнообразие материала для эволюционного процесса без снижения жизнеспособности особей; 2) расширяются возможности приспособления организмов к изменяющимся условиям среды и тем самым обеспечивается выживание группы организмов (популяции, вида) в целом. Комбинативная изменчивость используется в селекции с целью получения более ценного в хозяйственном отношении сочетания наследственных признаков. В частности применяется явление гетерозиса, повышения жизнеспособности, интенсивности роста и других показателей при гибридизации между представителями различных подвидов или сортов. Ярко выражено оно, например, у кукурузы (рис. 78), обусловливая значительный экономический эффект. Противоположный эффект даёт явление инбридинга или близкородственного скрещивания - скрещивания организмов, имеющих общих предков. Общность происхождения скрещиваемых организмов увеличивает у них вероятность наличия одних и тех же аллелей любых генов, а следовательно - вероятность появления гомозиготных организмов. Наибольшая степень инбридинга достигается при самоопылении у растений и самооплодотворении у животных. Гомозиготность увеличивает возможность проявления рецессивных аллельных генов, мутагенные изменения которых приводят к появлению организмов с наследственными аномалиями. Результаты изучения явления комбинативной изменчивости используются в медико-генетическом консультировании, особенно на его втором и третьем этапах: прогноз потомства, формирование заключения и объяснение смысла генетического риска. В консультировании будущих супружеских пар используется установление вероятности наличия у каждого из двух индивидуумов аллелей, полученных от общего предка и идентичных по происхождению. Для этого используют коэффициент родства, выражаемый в долях единицы. У монозиготных близнецов он равен 1, у родителей и детей, братьев и сестёр - 1/2, у деда и внука, дяди и племянника - 1/4, у двоюродных сибсов (братьев и сестёр) - 1/8, у троюродных сибсов - 1/32 и т.д.
Примеры: У цветка ночная красавица есть ген красного цвета лепестков А, и ген белого цвета а. Организм Аа имеет розовый цвет лепестков. Таким образом, у ночной красавицы нет гена розового цвета, розовый цвет возникает при сочетании (комбинации) красного и белого гена.
У человека есть наследственное заболевание серповидноклеточная анемия. АА – норма, аа – смерть, Аа – СКА. При СКА человек не может переносить повышенных физических нагрузок, при этом он не болеет малярией, т.е. возбудитель малярии малярийный плазмодий не может питаться неправильным гемоглобином. Такой признак полезен в экваториальном поясе; для него нет гена, он возникает при сочетании генов А и а.
Типы неаллельных взаимодействий: эпистаз доминантный и рецессивный
Неалле́льные ге́ны - это гены, расположенные в различных участках хромосом и кодирующие неодинаковые белки. Неаллельные гены также могут взаимодействовать между собой.
При этом либо один ген обусловливает развитие нескольких признаков, либо, наоборот, один признак проявляется под действием совокупности нескольких генов. Выделяют три формы и взаимодействия неаллельных генов:
комплементарность;